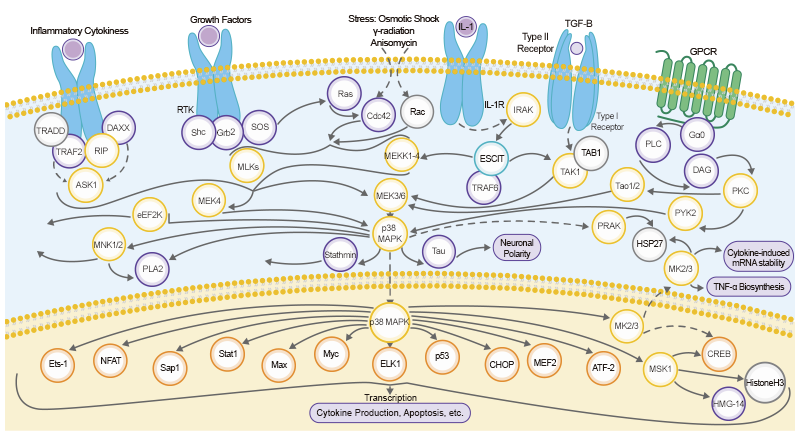
Reaching the golden years doesn’t always feel so golden. As we age, disease, injury, and other stress factors from the environment will damage our bodies' cells. Most cells may be able to repair that damage, while our immune system usually clears those damaged cells through a process called apoptosis.[1] However, if cellular repair and clearance is not effective, the residual damaged cells will further weaken the immune system and deteriorate other biological processes. Is there a possibility that we can avoid this cellular damage and improve the health of older people? A cellular state known as senescence might hold the key to this question.[1, 2] During senescence, the damaged cells irreversibly stop dividing and resist being removed. [3] Researchers have shown that determining senescence biomarkers could lead to new therapies for the inflammatory disease caused by senescence in older people.[4]